
Adaptive Evolution
Section Goals
By the end of this section, you will be able to do the following:
- Explain the different ways natural selection can shape populations
- Describe how these different forces can lead to different outcomes in terms of the population variation
Biological evolution, simply put, is descent with modification.
Biological evolution is not simply a matter of change over time. Lots of things change over time: trees lose their leaves and mountain ranges rise and erode, but they aren’t examples of biological evolution because they don’t involve descent through genetic inheritance.
The central idea of biological evolution is that all life on Earth shares a common ancestor, just as you and your cousins share a common grandmother.
Through the process of descent with modification, the common ancestor of life on Earth gave rise to the fantastic diversity that we see documented in the fossil record and around us today. Evolution means that we’re all distant cousins: humans and oak trees; hummingbirds and whales.
Evolution occurs at different scales:
- Microevolution (small-scale evolution): Changes in gene frequency within a population (not resulting in speciation)
- Macroevolution (large-scale evolution): Changes in gene frequency that result in speciation (one population is different enough from other populations that it is no longer the same species)
Although the term “evolution” is often used synonymously with “natural selection,” they are actually referring to different concepts. Evolution is an observable phenomenon in which gene frequencies change over time, but it does not explain why a population is undergoing evolution. This is where natural selection—and other mechanisms—come into play. They explain “the why.”
Natural Selection
Natural selection acts on the population’s heritable traits: selecting for beneficial alleles that allow for environmental adaptation, thus increasing their frequency in the population, while selecting against deleterious alleles and thereby decreasing their frequency. Scientists call this process adaptive evolution. Natural selection acts on entire organisms, not on an individual allele within the organism. An individual may carry a very beneficial genotype with a resulting phenotype that, for example, increases the ability to reproduce (fecundity), but if that same individual also carries an allele that results in a fatal childhood disease, that fecundity phenotype will not pass to the next generation because the individual will not live to reach reproductive age. Natural selection acts at the individual’s level. It selects for individuals with greater contributions to the gene pool of the next generation. Scientists call this an organism’s evolutionary (Darwinian) fitness.
Fitness is often quantifiable and is measured by scientists in the field. However, it is not an individual’s absolute fitness that counts, but rather how it compares to the other organisms in the population. Scientists call this concept relative fitness, which allows researchers to determine which individuals are contributing additional offspring to the next generation, and thus, how the population might evolve.
There are several ways selection can affect population variation: stabilizing selection, directional selection, diversifying selection, frequency-dependent selection, and sexual selection. As natural selection influences the allele frequencies in a population, individuals can either become more or less genetically similar and the phenotypes can become more similar or more disparate.
Stabilizing Selection
If natural selection favors an average phenotype, selecting against extreme variation, the population will undergo stabilizing selection (Figure 1). In a mouse population that lives in the woods, for example, natural selection is likely to favor mice that best blend in with the forest floor and are less likely for predators to spot. Assuming the ground is a fairly consistent shade of brown, those mice whose fur is most closely matched to that color will be most likely to survive and reproduce, passing on their genes for their brown coat. Mice that carry alleles that make them a bit lighter or a bit darker will stand out against the ground and be more likely to fall victim to predation. As a result of this selection, the population’s genetic variability will decrease.
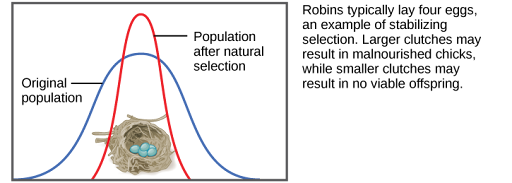
Directional Selection
When the environment changes, populations will often undergo directional selection (Figure 2), which selects for phenotypes at one end of the spectrum of existing variation. A classic example of this type of selection is the evolution of the peppered moth in eighteenth- and nineteenth-century England. Prior to the Industrial Revolution, the moths were predominately light in color, which allowed them to blend in with the light-colored trees and lichens in their environment. However, as soot began spewing from factories, the trees darkened, and the light-colored moths became easier for predatory birds to spot. Over time, the frequency of the moth’s melanic form increased because they had a higher survival rate in habitats affected by air pollution because their darker coloration blended with the sooty trees. Similarly, the hypothetical mouse population may evolve to take on a different coloration if something were to cause the forest floor where they live to change color. The result of this type of selection is a shift in the population’s genetic variability toward the new, fit phenotype.
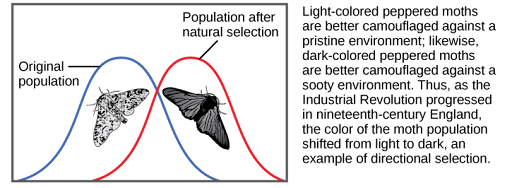
Diversifying Selection
Sometimes two or more distinct phenotypes can each have their advantages for natural selection, while the intermediate phenotypes are, on average, less fit. Scientists call this diversifying selection (Figure 3) We see this in many animal populations that have multiple male forms. Large, dominant alpha males use brute force to obtain mates, while small males can sneak in for furtive copulations with the females in an alpha male’s territory. In this case, both the alpha males and the “sneaking” males will be selected for, but medium-sized males, who can’t overtake the alpha males and are too big to sneak copulations, are selected against. Diversifying selection can also occur when environmental changes favor individuals on either end of the phenotypic spectrum. Imagine a mouse population living at the beach where there is light-colored sand interspersed with patches of tall grass. In this scenario, light-colored mice that blend in with the sand would be favored, as well as dark-colored mice that can hide in the grass. Medium-colored mice, alternatively, would not blend in with either the grass or the sand, and thus predators would most likely eat them. The result of this type of selection is increased genetic variability as the population becomes more diverse.
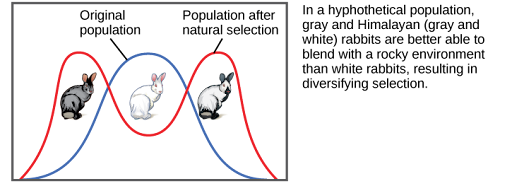
Different types of natural selection can impact the distribution of phenotypes within a population (refer back to Figures 1, 2, and 3). In recent years, factories have become cleaner, and less soot is released into the environment. What impact do you think this has had on the distribution of moth color in the population?
Answer:
Moths have shifted to a lighter color.
Sexual Selection
Darwin noticed that there were many traits and behaviors of organisms that could not just be explained by the ability to survive. For example, the brilliant plumage of peacocks should actually lower their rates of survival. That is, the peacocks’ feathers act like a neon sign to predators, advertising “Easy, delicious dinner here!” But if these bright feathers only lower peacocks’ chances of survival, why do they have them? The same can be said of similar characteristics of other animals, such as the large antlers of male stags or the wattles of roosters, which also seem to be unfavorable to survival. Again, if these traits only make the animals less likely to survive, why did they develop in the first place? And how have these animals continued to survive with these traits over thousands and thousands of years? Darwin’s answer to this conundrum was the theory of sexual selection: the evolution of characteristics, not because of survival advantage, but because of mating advantage.
Intrasexual Selection
Members of one sex compete against each other, and the winner mates with members of the opposite sex. It is “intra-sex” because the competition is occurring within one sex.
Intersexual Selection
If members of one sex are attracted to certain qualities in mates, then those desired qualities get passed on in greater numbers because their possessors mate more often. It is “inter-sex” because one sex affects selection in the other sex.
Did I Get It?
Sexual selection has shaped many extreme adaptations that help organisms find mates. Perhaps one of the most extreme examples is in Australian redback spiders– also known as Australian black widows (Figure 4). The venomous female redback spider poses a danger to humans, and to male redback spiders, which are often eaten by their mates. Males seem to go out of their way to make this happen, flipping themselves over and presenting their abdomens to the female while mating.

This behavior might at first seem like one that selection would act against. After all, how could risking one’s life be adaptive? Remember that evolutionary fitness is about getting genes into the next generation, not just survival. Perhaps this strange behavior is favored by sexual selection because it gives males a fitness boost.
Scientist Spotlight: Maydianne Andrade

Evolutionary ecologist Maydianne Andrade is known for her work researching the behavior of the cannibalistic Australian redback spiders. Listen to Canada’s national public radio interview with Andrade, including a discussion of spiders, unconscious bias, imposter syndrome, and the burden carried by Black scientists.
Frequency-Dependent Selection
Another type of selection, frequency-dependent selection, favors phenotypes that are either common (positive frequency-dependent selection) or rare (negative frequency-dependent selection). We can observe an interesting example of this type of selection in a unique group of Pacific Northwest lizards. Male common side-blotched lizards come in three throat-color patterns: orange, blue, and yellow. Each of these forms has a different reproductive strategy: orange males are the strongest and can fight other males for access to their females. Blue males are medium-sized and form strong pair bonds with their mates. Yellow males (Figure 5) are the smallest, and look a bit like females, which allows them to sneak copulations. Like a game of rock-paper-scissors, orange beats blue, blue beats yellow, and yellow beats orange in the competition for females. That is, the big, strong orange males can fight off the blue males to mate with the blue’s pair-bonded females, the blue males are successful at guarding their mates against yellow sneaker males, and the yellow males can sneak copulations from the potential mates of the large, polygynous orange males.

In this scenario, natural selection favors orange males when blue males dominate the population. Blue males will thrive when the population is mostly yellow males, and yellow males will be selected for when orange males are the most populous. As a result, populations of side-blotched lizards cycle in the distribution of these phenotypes—in one generation, orange might predominate, and then yellow males will begin to rise in frequency. Once yellow males comprise a majority of the population, blue males will be selected. Finally, when blue males become common, orange males once again will be favored.
Negative frequency-dependent selection serves to increase the population’s genetic variability by selecting for rare phenotypes; whereas, positive frequency-dependent selection usually decreases genetic variability by selecting for common phenotypes.
Did I Get It?
Adaptation
An adaptation is a feature that arose and was favored by natural selection for its current function. Adaptations help an organism survive and/or reproduce in its current environment.
Adaptations can take many forms: a behavior that allows better evasion of predators, a protein that functions better at body temperature, or an anatomical feature that allows the organism to access a valuable new resource — all of these might be adaptations. A platypus’s webbed feet are an adaptation for swimming. A snow leopard’s thick fur is an adaptation for living in the cold. A cheetah’s fast speed is an adaptation for catching prey.
Many of the things that impress us most in nature are thought to be adaptations.

Mimicry of leaves by insects is an adaptation for evading predators (Figure 6). The creosote bush is a desert-dwelling plant that produces toxins that prevent other plants from growing nearby, thus reducing competition for nutrients and water. Echolocation in bats is an adaptation for catching insects.
So what’s not an adaptation? The answer: a lot of things. One example is vestigial structures. A vestigial structure is a feature that was adaptive for the organism’s ancestor, but that evolved to be non-functional because the organism’s environment changed. Fish species that live in completely dark caves have vestigial, non-functional eyes. When their sighted ancestors ended up living in caves, there was no longer any natural selection that maintained the function of the fish’s eyes. So fish with better sight no longer out-competed fish with worse sight. Today, these fish still have eyes — but they are not functional and are not an adaptation; they are just the by-products of the fishes’ evolutionary history.
Misconceptions about natural selection
Because natural selection can produce amazing adaptations, it’s tempting to think of it as an all-powerful force, urging organisms on, constantly pushing them in the direction of progress — but this is not what natural selection is like at all.
Natural selection is not all-powerful; it does not produce perfection. It’s all about getting genes into the next generation, and if your genes are “good enough” to do that, you don’t have to be perfect. This should be clear just by looking around us: human populations carry genes that cause disease, plants may not have the genes to survive a drought, and a predator may not be quite fast enough to catch her prey every time she is hungry. No population or organism is perfectly adapted.
It’s more accurate to think of natural selection as a process rather than as a guiding hand. Natural selection is the simple result of variation, differential reproduction, and heredity — it is mindless and mechanistic. It has no goals; it’s not striving to produce “progress” or a balanced ecosystem.


This is why “need,” “try,” and “want” are not very accurate words when it comes to explaining evolution. The population or individual does not “want” or “try” to evolve, and natural selection cannot try to supply what an organism “needs.” Natural selection can only select from whatever variation already exists in the population. It does not create anything from scratch. Thus, it is limited by a population’s existing genetic variability and whatever new alleles arise through mutation and gene flow.
Natural selection is also limited because it works at the individual, not allele level, and some alleles are linked due to their physical proximity in the genome, making them more likely to pass on together (linkage disequilibrium). Any given individual may carry some beneficial and some unfavorable alleles. It is the alleles’ net effect, or the organism’s fitness, upon which natural selection can act. As a result, good alleles can be lost if individuals who carry them also have several overwhelmingly bad alleles. Likewise, bad alleles can be kept if individuals who have enough good alleles to result in an overall fitness benefit carry them.
Read more about how natural selection does not produce perfectly engineered traits or perfect populations (i.e., populations with no deleterious genes).
At the opposite end of the scale, natural selection is sometimes interpreted as a completely random process. This is also a misconception. The genetic variation that occurs in a population because of mutation is random — but selection acts on that variation in a very non-random way: genetic variants that aid survival and reproduction are much more likely to become common than variants that don’t. Natural selection is NOT random!
Finally, it is important to understand that not all evolution is adaptive. While natural selection selects the fittest individuals and often results in a more fit population overall, other forces of evolution, including genetic drift and gene flow, often do the opposite: introducing deleterious alleles to the population’s gene pool. Evolution has no purpose—it is not changing a population into a preconceived ideal. It is simply the sum of the various forces that we have described in this chapter and how they influence the population’s genetic and phenotypic variability.
CC Licensed Content, Shared Previously, Included in Mechanisms of Evolution
- Biology 2e. Authors: Mary Ann Clark, Matthew Douglas and Jung Choi. Provided by: OpenStax CNX. Located at: Biology 2e. License: CC BY: Attribution 4.0.
- Biology for Majors II. Authors: Shelly Carter and Monisha Scott. Provided by: Lumen Learning. Located at: Biology for Majors II | Simple Book Production. License: CC BY: Attribution 4.0.
- An Interactive Introduction to Organismal and Molecular Biology, 2nd ed. Author: Andrea Bierema. Provided by: Pressbooks. Located at: An Interactive Introduction to Organismal and Molecular Biology, 2nd ed. – Simple Book Publishing. License: CC BY-NC-SA 4.0
- Misconceptions about Natural Selection. Provided by: UC Museum of Paleontology Understanding Evolution. Located at: https://evolution.berkeley.edu/evolution-101/mechanisms-the-processes-of-evolution/misconceptions-about-natural-selection License: CC BY-NC-SA 4.0
- Sexual Selection. Provided by: UC Museum of Paleontology Understanding Evolution. Located at: https://evolution.berkeley.edu/evolution-101/mechanisms-the-processes-of-evolution/sexual-selection/. License: CC BY-NC-SA 4.0
