
Meiosis
In organisms that replicate sexually, in each new generation genetic information is remixed and recombined from two parental cells. This happens when two parental gametes – each of which contain half of a genome – fuse together.
A cell about to undergo meiosis is called a meiocyte. Meiosis is mechanistically like mitosis but involves two rounds of cell division. In meiosis I, homologous chromosome pairs separate and are distributed among daughter cells. Note: this process is often described as segregation of the homologs. This word has a very negative meaning in a modern social context, referring primarily to segregation of ethnic or racial groups of people. In a biological context, it means a physical separation and compartmentalization. It is a holdover from work like Gregor Mendel’s in the 1860’s. He used the word to describe of the equal segregation of dominant and recessive characters among offspring.
Meiosis I is the reductional stage of mitosis since it reduces the total number of chromosomes. In meiosis II, sister chromatids separate, just as in mitosis. Meiosis II is an equatorial division just like mitosis, since it does not change chromosome number. Meiosis I and II both follow the same general sequence of events as mitosis: prophase, metaphase, anaphase, and telophase. In nomenclature, these stages are distinguished from one another by the addition of the Roman numerals I or II. For example, “prophase”, with no Roman numeral, indicates the stage of mitosis. “Prophase I” indicates prophase of meiosis I, and “prophase II” indicates prophase of meiosis II. Meiosis I and II include two rounds of cytokinesis as well: for each parent meiocyte, there are thus four daughter cells produced. An overview of the processes of Meiosis I and Meiosis II is in Figure 9.
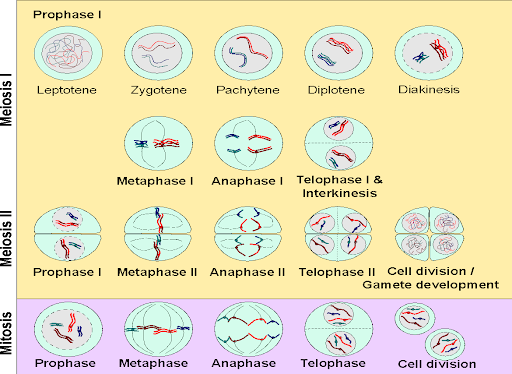
The biggest difference between Prophase I and mitotic Prophase is that, during Prophase I, homologous chromosomes synapse, or pair, forming a synaptonemal complex. This holds the homologs together and is important for the proper separation of chromosomes, ensuring that exactly one chromosome from each pair is destined for each daughter cell. It is also important for the exchange of genetic information between the homologous pairs, called crossing over or meiotic recombination.
The condensation of chromosomes and formation of the synaptonemal complex can be observed microscopically, and so Prophase I can be further divided based on those events. Leptotene is the first stage after interphase, when chromosomes begin to condense. Next is the zygotene stage, when homologous pairs come together to form the synaptonemal complex. During the pachytene stage, the chromosomes are paired all along their length, and there is crossing over of genetic information between the homologous chromosomes. The pairs begin to partially separate during diplotene, making crossovers visible in the space between the pairs. The nuclear envelope dissolves during diakinesis.
Because genetic information is exchanged reciprocally between parental chromosomes, crossing over (Figure 10) means that chromosomes do not have to be inherited intact from generation to generation, but it does mean that no genetic information is lost. Crossing over thus plays an important role in increasing genetic diversity while maintaining genomic integrity. Although only one crossover is shown per chromosome in Figure 10, multiple crossovers can and do occur between homologs. The daughter chromosome could be a patchwork sequence from one or the other parental chromosome.
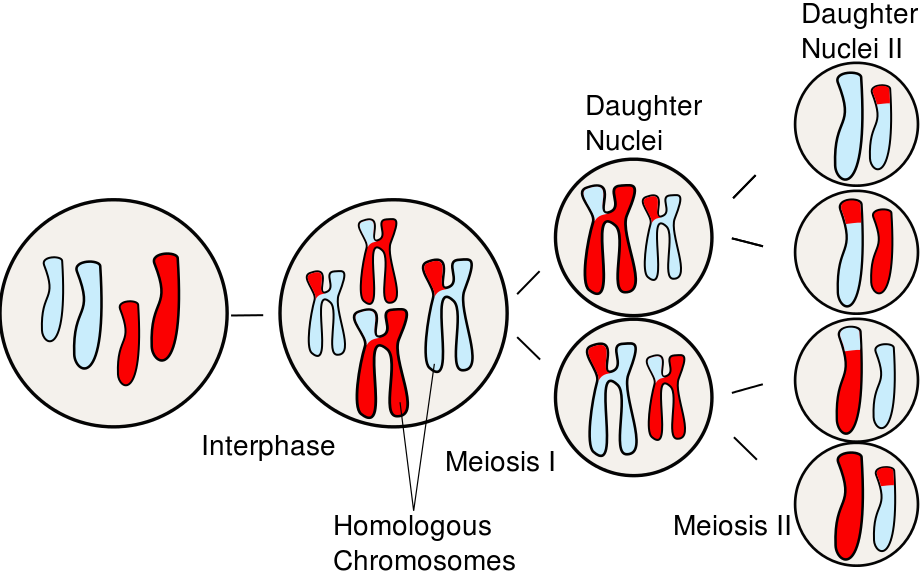
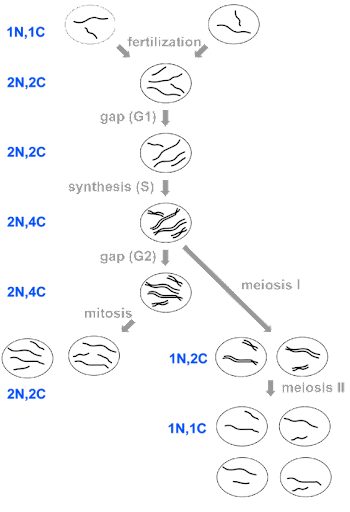
Following the diakinesis stage is metaphase I. As in the metaphase of mitosis, the chromosomes align on the equatorial plate of the cell. However, because the homologous chromosomes are paired, they orient in a way that chromosome pairs (rather than sister chromatids) are destined to be partitioned. The alignment of the pairs is what determines the combination of alleles in the daughter cells. With the transition to anaphase I, the homologous pairs separate and migrate toward their respective poles of the cell. During telophase I, nuclei reform and cytokinesis may separate the two cells into daughters. Because only one chromosome from each pair is distributed to each daughter cell, the daughter cells are haploid (1n, 2C) as they enter meiosis II.
The stages of meiosis II are much like mitosis. Chromosomes condense in prophase II and align on the equatorial plate during metaphase II. Chromatids separate and segregate during anaphase II, and nuclei reform and the cells separate during telophase II and cytokinesis. At the end of meiosis II, the DNA content of the daughter cells is 1n, 2C.
The stages of meiosis and mitosis are summarized in Figure 11, with DNA content for each stage indicated.
As in mitosis, checkpoints during meiosis ensure that each step is completed before the cell divides. Meiotic checkpoints monitor for synapse formation and resolution of crossovers, preventing cycle progression if these steps are not completed appropriately[1].

Interestingly, these checkpoints may affect fertility in hybrid offspring from different species. For example, a mule is a hybrid of a horse (2n = 64) and a donkey (2n = 62), with 63 chromosomes. The odd number of chromosomes cannot pair during meiosis I, and the failure of synapse leads to a block to gametogenesis in mid-meiosis[2].
Media Attributions
- The stages of meiosis I and II © Open Genetics Lectures, Chapter 15 is licensed under a CC BY-NC (Attribution NonCommercial) license
- Subramanian, V. V. & Hochwagen, A. The Meiotic Checkpoint Network: Step-by-Step through Meiotic Prophase. Cold Spring Harb. Perspect. Biol. 6, a016675 (2014). ↵
- Han, H. et al. Testicular Characteristics and the Block to Spermatogenesis in Mature Hinny. Asian-Australas. J. Anim. Sci. 29, 793–800 (2016). ↵

{kind=link}