
Chromosomes and sex development
Sex is a phenotype, like any other observable trait. Sex determination genes are genes that control the development of sex-associated traits.
In mammals, fruit flies, and some flowering plants embryos, females usually (but not always) have an XX genotype while males usually (but not always) have an XY genotype. In birds and some reptiles and amphibians, males typically have a ZZ genotype and females have ZW genotype. Note that females are not always the homogametic sex – they don’t always have two of the same chromosome.
These genotypes are associated with either male or female phenotypes due to the presence of sex determination genes located on the sex chromosomes. But the mechanism for development of sex-associated phenotypes is different depending on species. For example, although humans and Drosophila both have X and Y sex chromosomes, they have different mechanisms for determining sex.
Table 2 lists chromosomal methods of sex determination in several species. In humans and other mammals, the SRY gene on the Y chromosome triggers the development of testes in the early embryo.
Fruit flies and certain other insects also use an XY system of sex determination, but the mechanism of sex determination is different. For fruit flies, the ratio of X chromosomes to autosomes determines sex phenotype, due to the expression of autosomal genes that in turn influences the expression of sex-determination genes on the X chromosome.
In other insects, there’s no Y chromosome at all! The number of X chromosomes influences maleness. And in birds and some other species, the DMRT1 gene on the Z chromosome initiates the process of sex development, but it is haploinsufficient: one copy of the gene does not produce enough gene product to trigger maleness, so ZW individuals will typically develop female anatomy.
Note: some species do not use sex chromosomes! Sex in some species can be determined by autosomes or even environmental conditions. In honeybees, sex is determined by whether eggs are fertilized: unfertilized eggs develop into males, while fertilized eggs develop into females. In many turtles, sex is determined by environmental temperature: cooler temperatures are associated with male development, and warmer temperatures with female development[1].
| Organism | Human and other mammals
|
Drosophila (fruit fly) | Some insects and other organisms | Birds, reptiles, some fishes and amphibians |
|---|---|---|---|---|
| Chromosomal System | XX-XY | XX-XY (XA system) | XX-XO | ZZ-ZW |
| Chromosomal method of determination | Presence of Y chromosome triggers maleness during fetal development
|
Ratio of X:autosomal chromosomes determines sex
|
Number of X chromosomes determines sex; there is no Y chromosome. | Two Z chromosomes required for maleness |
| Mechanism | SRY gene on the Y chromosome sets off a series of events leading to development of testes. (Other genes both autosomal and sex-linked are also involved) | Expression of sex-determining genes on the X-chromosome is affected by timing (and level) of expression of autosomal genes
|
May vary among species: In C. elegans is related to the amount of an X-encoded protein which can inhibit maleness | DMRT1 gene on the Z chromosome is haploinsufficient; one copy of the gene can’t produce enough protein to trigger male development |
In all organisms, the sex chromosomes are associated with sex determination because they house some, but not all, of the genes responsible for determining sex. (Other sex-determination genes are located on the autosomes, as discussed below.). The sex chromosomes also house genes not responsible for sex determination at all. These are called sex-linked genes, but they have nothing to do with sex beyond their chromosomal locus. Some examples in humans are a gene linked with color-blindness and a gene that affects the strength of tooth enamel.
Test Your Understanding
Sex differentiation during human development
So, given what we know about genes, chromosomes, and gene expression, why does the presence of a Y chromosome typically trigger maleness during human development? The answer to that involves a network of both sex-linked and autosomal genes.
Sex is a phenotype. Typically, in most sexually dimorphic species, there are multiple characteristics, in addition to sex organs, that distinguish male from female individuals. Those sex-associated traits are phenotypes just like hair color or eye color or wing shape. Those phenotypes can be genetically (or in some cases environmentally) determined.
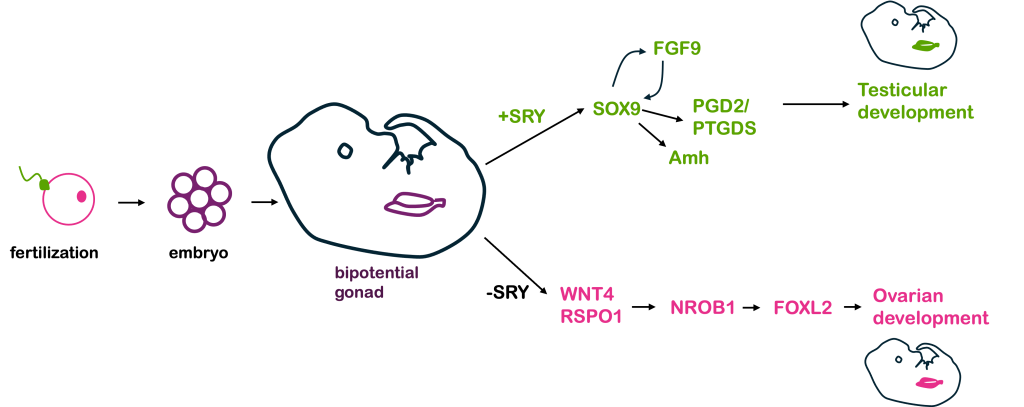
In humans and other mammals, the Y chromosome carries the SRY gene. The SRY protein encoded by the gene is a transcription factor. In early stages of human development, a human embryo develops a bi-potential genital ridge – that is, tissue that has the potential to become either ovaries or testes. Early embryos also have two systems of ducts, Wolffian and Müllerian, which can develop into the male and female reproductive tracts, respectively.
If a Y chromosome is present, the transcription factor SRY is produced. In turn, SRY activates the expression of Sox9, which is also a transcription factor. Sox9 in turn activates other genes, which eventually lead to the development of testes.
The testes in turn produce testosterone and anti-Müllerian hormone, or AMH. Testosterone (and other hormones, including 5α-dihydrotestosterone) trigger formation of other organs in the male reproductive system from the Wolffian duct tissue, while AMH causes degeneration of the Müllerian duct and suppresses the development of female sex structures.
In the absence of SRY (as in individuals with an XX genotype), an alternative set of molecular signals is typically activated, including WNT4, RSPO1, DHH (Desert Hedgehog), and β-catenin. These lead to development of ovaries. The ovaries then produce estrogen and trigger development of the uterus, oviducts, and cervix from the Müllerian duct.
The sequential activation of these genes is illustrated in Figure 7, below.
Thus, in humans and other mammals, the SRY protein product sets into motion this cascade of events that leads to maleness. However, SRY is not the only gene involved in this process – and, in fact, all the other genes shown in Figure 7 are autosomal, not sex-linked! A loss of function in any of the autosomal genes will disrupt the sex determination pathway, leading to differences in sex development, discussed in greater detail below.
Differences of sex development in humans
Although the term “biological sex” is used quite often, this term is an oversimplification of the biology underlying the development of sex characteristics. There are a number of ways males and females differ from one another. As a result, there are many ways “biological sex” might be defined.
In this module, we’ve so far been focused on chromosomal sex, where an XY individual is considered male and an XX individual is considered female. But sex is usually determined or assigned at birth based on the visible presence of a penis or a vulva in a newborn. Sex can be determined by genitalia even before birth, when genitals are clearly visible by ultrasound.
Genitalia define anatomical sex. This is also sometimes called phenotypic sex, although this is an oversimplification of term phenotype. Gonadal sex refers to the presence of either testes or ovaries. Although individuals show a range of hormonal levels, human males and females tend to have different ranges of androgens (males typically have more, females typically have less) and estrogens (females typically have more, males typically have less). This is hormonal sex.
At puberty, both males and females develop secondary sex characteristics. In males, this includes deepening of the voice, growth of facial and body hair, and broadening of the shoulders. In females, secondary sex characteristics include growth of breast tissue, widening of hips, and onset of menses. These secondary sex characteristics typically (though not always) develop according to hormonal sex.
You might expect that these ways of defining biological sex (chromosomal, anatomical, gonadal, and hormonal) all align: XY individuals would have a penis, have testes, have relatively high levels of androgens like testosterone and low levels of estrogens, and develop male secondary sex characteristics at puberty. But this is not always true: it is possible to have an XY genotype but have female genitalia, gonads, and secondary sex characteristics. It is possible to have an XX genotype and have male genitalia, gonads, and secondary sex characteristics. It is possible to have female external genitalia and internal testes. It is possible to have genitalia with both male and female characteristics, or indeterminate characteristics. In fact, it is possible to have almost any combination of chromosomal, genital, gonadal, and hormonal sex.
Genetically, these sex phenotypes are due to genotypic changes in any one of the networks of genes responsible for sex determination and differentiation – and because there are so many genes involved in the process, there are many Differences of Sex Development (DSDs) in the human population. Another term for DSD is intersex. Some examples of DSDs in humans are listed below, although this is not an exhaustive list.
Sex chromosome aneuploidy. Remember that aneuploidy refers to an atypical number of chromosomes. They often result from nondisjunction during meiosis or mitosis: either homologous pairs or sister chromatids fail to separate during anaphase. Aneuploidies of autosomes are relatively rare in the human population, since additional copies – or too few copies – of most chromosomes would have lethal effects on a developing embryo. (Some exceptions are Trisomy 21 – Down syndrome, Trisomy 13, and Trisomy 18, all of which have significant phenotypic effects and are linked with shortened lifespan.) Aneuploidies of sex chromosomes, however, are by far more common. This is likely because the Y chromosome has very few genes – none required for life – and any extra copies of the X chromosome can be inactivated (discussed more in the next section).
Sex chromosome aneuploidies are the most common DSDs in the human population as well as the most common aneuploidies in the human population. Humans with sex chromosome aneuploidies usually have a normal lifespan but may be infertile, may have neurological and/or cognitive disabilities, or they may have few or no measurable phenotypic differences. It’s estimated that 50-75% of people with sex chromosome abnormalities never even know they have a sex chromosome aneuploidy.
Individuals without a Y chromosome typically – but not always – show a female phenotype, regardless of the total number of sex chromosomes. This is because the SRY gene on the Y chromosome is needed to initiate development of male reproductive structures. Individuals with one or more Y chromosomes typically – but not always – show a male phenotype since the presence of SRY triggers development of male reproductive structures during embryogenesis.
A selected list of human sex chromosome abnormalities is listed in Table 3: Sex chromosome aneuploidies in humans, below, with associated phenotype.
| Genotype | Phenotype |
| XO (One sex chromosome only) | Turner syndrome. Anatomical females present with symptoms that vary in severity but may include short stature, ovarian failure, cardiac defects, and/or infertility that may be corrected with fertility treatment. |
| XXY | Klinefelter syndrome. Anatomical males present with signs and symptoms that vary in severity but may include taller than average stature, weak bones, delayed puberty, decreased muscle mass, and low sex drive. Some may go undiagnosed. |
| XYY | XYY syndrome. Anatomical males with normal fertility and sexual development. May be taller than average, have increased risk of cystic acne, ADHD, and, to a lesser extent, autism spectrum disorder. May go undiagnosed. |
| XXX | Triple X syndrome. Anatomical females who are generally taller than average, some with subtle physical differences including wide-spaced eyes. Some patients may have learning disabilities and medical problems including infertility due to premature ovarian failure, but others may have mild or no symptoms and go undiagnosed. |
Translocation of SRY. In some anatomical males with an XX genotype, the SRY gene has been translocated to one of the X chromosomes or an autosome. This is often the result of an aberrant recombination event – crossing over between the X and Y chromosomes (or the Y and an autosome) during meiosis in the father. Such individuals appear phenotypically male but may have small testes or a urethra opening on the underside of the penis. Affected individuals may be shorter than average, require hormone treatments at puberty to trigger development of secondary sex characteristics, and may be infertile, but are otherwise healthy.
Swiyer syndrome. XY individuals who are phenotypically female, with functional vagina, uterus, and fallopian tubes but lacking ovaries. Because they lack ovaries, women with Swiyer syndrome are usually diagnosed in adolescence when they do not undergo a first period. Secondary sex characteristics do not develop without hormonal supplementation, but women can become pregnant with donated embryos. Women with Swiyer syndrome often have a deletion on the Y chromosome that inactivates the SRY gene, but the syndrome is also associated with mutations in the gene NROB1 on the X chromosome and several autosomal genes as well.
Complete androgen insensitivity (CAIS). 46, XY female phenotype. Mutations in the androgen receptor gene prevent cells of a developing embryo from responding to androgens, including testosterone. Although people with CAIS have normal-appearing female external genitalia and secondary sex characteristics, they have internal, undescended testes, and usually have a shortened vagina and lack a uterus.
Congenital adrenal hyperplasia (CAH). Mutations in the enzyme 21-hydroxylase block one step of cortisol synthesis. This in turn leads to a build-up of testosterone since cortisol and testosterone share parts of their biosynthetic pathway. This causes masculinization of the external genitalia in XX individuals, who may have ambiguous genitalia or even the appearance of normal male external genitalia. Such XX individuals have female gonads and internal genitalia.
5-alpha-reductase deficiency. These XY individuals have a loss of function mutation in the enzyme that converts testosterone to dihydrotestosterone (DHT). DHT is the hormone required for the development of external male genitalia during fetal development. Children with 5-alpha-reductase deficiency may appear to have female external genitalia or ambiguous external genitalia at birth. However, they have internal testes, and, at puberty, the testes produce much higher levels of testosterone that cause penile and scrotal tissue to grow and male secondary sex characteristics to develop. This particular DSD occurs at high frequency in certain communities in the Dominican Republic, where such people are called “guavedoces”. The term “guavedoces” very loosely translates to “penis at age twelve”, since children assumed to be female appear to grow a penis.
With this expanded understanding of biological sex, it becomes apparent that while individuals with unambiguously male or unambiguously female characteristics make up the majority of the human population, individuals frequently do not completely align with either category. Thus, although sex is often considered binary – male or female – this is not an accurate picture of human biology, since there are many ways differences in sex development can arise. Humans may be chromosomally male but phenotypically female, or vice versa. They may be chromosomally male, hormonally male, but phenotypically female. Or they may have nearly any other combination of chromosomal, anatomical, gonadal, and hormonal sex.
Estimates of how many individuals in the human population have DSDs range depending on who is counted. If only individuals with differences in external genitalia are counted, such individuals make up about 1/4500-1/2000 of the human population. However, DSDs may result in typical external genitalia but atypical alignment of chromosomal, hormonal, or gonadal sex. They may also include individuals with atypical numbers of sex chromosomes – many of whom may not even know they have such a condition. If individuals with atypical sex chromosome ploidy are included, some estimates are that individuals with DSDs make up about 2% of the human population. For comparison, this is about the same frequency as the red hair phenotype worldwide – not as uncommon as one might think. If you know people with red hair, you likely know someone with a difference of sex development, too, making sex not very binary at all.
Test Your Understanding
Media Attributions
- Selected genes required for sex © Amanda Simons is licensed under a CC BY-SA (Attribution ShareAlike) license
- US Department of Commerce, N. O. and A. A. What causes a sea turtle to be born male or female? https://oceanservice.noaa.gov/facts/temperature-dependent.html. ↵

{kind=link}